

Друзья, я не большой специалист в физиологии и других “логиях”, поэтому не претендую на истину в последней инстанции.
Мне хотелось бы немного рассказать о том, что, как мне кажется, важно знать альпинистам и скалолазам.
Сегодня речь пойдет о мышцах.
Строение скелетных мышц (СМ).
СМ состоит из многочисленных удлиненных волокон (мышечных волокон, МВ). Каждое МВ представляет собой многоядерную клетку, покрытую эластичной оболочкой – сарколеммой.
Двигательные нервы входят в различных точках в МВ и передают ему электрический импульс, вызывающий сокращение.
И так, МВ имеет оболочку — сарколемму. Под оболочкой находится полужидкая среда — саркоплазма, в которой, по длине всего волокна, расположены в форме пучков нитевидные образования — миофибриллы (МФ). Они занимают практически все пространство ограниченное сарколеммой.
Увеличение количества МФ в мышечном волокне приводит к увеличению его силы и размера.
В саркоплазме МВ обнаруживаются и ряд других структур: митохондрии (МХ), рибосомы, трубочки и цистерны саркоплазматической сети (саркоплазматический ретикулум, СПР), различные вакуоли, «капельки» гликогена и жиров, играющие роль топлива для работающей мышцы.
В МВ СПР образует цистерны, где скапливаются ионы кальция. Для мышечного сокращения принципиальное значение имеет скорость откачивания ионов кальция из саркоплазмы, что, как мы увидим дальше, приводит к расслаблению мышцы.
Каждая МФ состоит из последовательно расположенных повторяющихся элементов — саркомеров. Границами саркомера служат т. н. Z-линии.
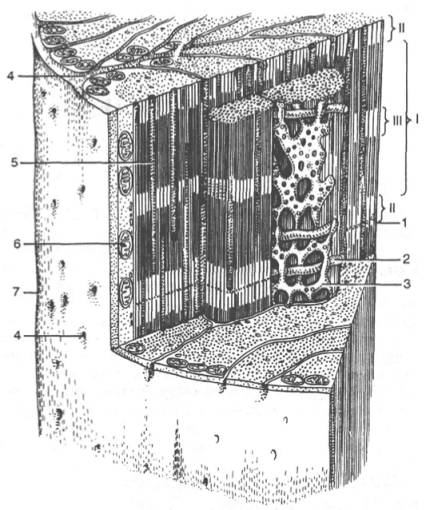
1-Z-линия; 2-Т-система; 3-саркоплазматическа сеть; 4-устье Т-системы; 5-гликоген; 6-митохондрия; 7-сарколемма; I-A-диск; II-I-диск; III-H-зона;
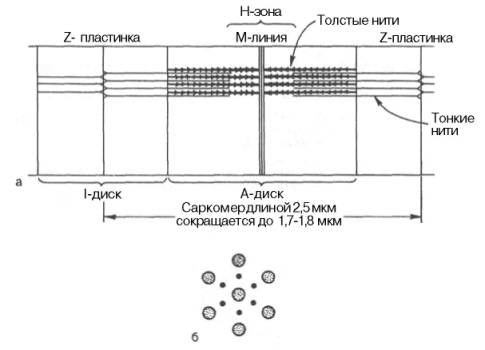
а-схематичное изображение структуры саркомера; б-расположение толстых(миозиновых) и тонких (актиновых) нитей (поперечное сечение).
Миозин составляет 50-55% от сухой массы МФ. Миозин обладает т. н. АТФазной активностью, т. е. способностью катализировать расщепление АТФ на АДФ и фосфорную к-ту, Н3РО4.
Молекула миозина имеет сильно вытянутую форму и состоит из нескольких субъединиц — две тяжелые полипептидные цепи, закрученные в спираль и несколько легких цепей, которые совместно с концами тяжелых цепей образуют глобулу («головка» миозина), которая способна соединяться с актином. Эти «головки» выдаются из основного стержня молекулы. И именно эти участки миозина и проявляют АТФазную активность.
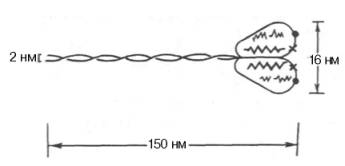
Строение молекулы мозина
Актин составляет 20% сухой массы МФ.
Известны две формы актина. Глобулярный G-актин и фибриллярный F-актин. Молекула G-актина состоит из одной полипептидный цепочки. При повышении ионной силы до физиологического уровня G-актин полимеризуется в F-актин. На электронных микрофотографиях он выглядит как две нити бус, закрученных одна вокруг другой.

1-актин; 2-тропомиозин; 3-тропонин С; 4-тропонин I; 5-тропонин T;
Кроме актина и миозина большое значение имеют белки тропомиозин и тропонин. Эти два белка образуют комплекс, называемый нативным тропомиозином. Он прикрепляется к актиновым филаментам и придает актомиозину (комплекс актина и миозина) чувствительность к ионам кальция.
Так же в мышцах содержится ряд важных небелковых веществ: АТФ, АДФ, АМФ, креатинфосфат, креатин, креатинин, свободные аминокислоты, гликоген, липиды(жиры) и др.
Одна из важнейших функций креатинфосфата – ресинтез АТФ, которая тратится при работе мышцы. При этом креатинфосфат превращается в креатин и фосфат. Эти молекулы распространяются по саркоплазме и активируют ферментативную активность связанную с синтезом АТФ.
Кроме того, предполагается что свободный креатин оказывает возбуждающие действие на белковый синтез в скелетных мышцах. Показано, что между содержанием сократительных белков и содержанием креатина имеется строгое соответствие. Свободный креатин, видимо, влияет на синтез и-РНК, т. е. на транскрипцию в ядрышках МВ, либо активирует деятельность ядерных митохондрий, которые начинают в большей мере вырабатывать АТФ, которая используется для транскрипции ДНК.
Механизм мышечного сокращения.
В настоящее время принято считать, что биохимический цикл мышечного сокращения состоит из 5 стадий:
1) Миозиновая «головка» может гидролизовать АТФ до АДФ и Н3РО4 (Pi), но не обеспечивает освобождения продуктов гидролиза (см. рис. а);
2) Содержащая АДФ и Н3РО4 миозиновая «головка» может свободно вращаться под большим углом и (при достижении нужного положения) связываться с F-актином, образуя с осью фибриллы угол около 90° (см. рис. б);
3) Это взаимодействие обеспечивает высвобождение АДФ и Н3РО4 из актин-миозинового комплекса. Актомиозиновая связь имеет наименьшую энергию при величине угла 45°, поэтому изменяется угол миозина с осью фибриллы с 90° на 45° (примерно) и происходит продвижение актина (на 10–15 нм) в направлении центра саркомера (см. рис. в);
4) новая молекула АТФ связывается с комплексом миозин–F-актин (см. рис. г);
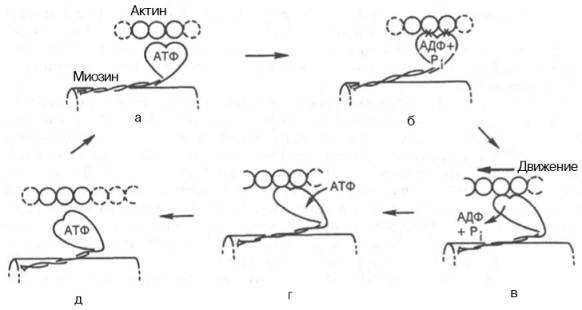
5) Комплекс миозин–АТФ обладает низким сродством к актину, и поэтому происходит отделение миозиновой (АТФ) «головки» от F-актина. Последняя стадия и есть собственно расслабление, которое отчетливо зависит от связывания АТФ с актин-миозиновым комплексом (см. рис. д). Затем цикл возобновляется.
Регуляция сокращения и расслабления мышц.
Ключевая регуляторная роль принадлежит ионам Са2+. Установлено, что МФ обладают способностью взаимодействовать с АТФ и сокращаться в его присутствии лишь при наличии в среде определенных концентраций ионов кальция.
Наибольшая сократительная активность наблюдается при концентрации ионов Са2+ около 10–6–10–5 М.
При понижении концентрации до 10–7 М или ниже мышечные волокна теряют способность к укорочению и развитию напряжения в присутствии АТФ.
По современным представлениям, в покоящейся мышце (в МФ и межфибриллярном пространстве) концентрация ионов Са2+ поддерживается ниже пороговой величины в результате связывания их структурами (трубочками и пузырьками) саркоплазматической сети и так называемой Т-системой.
Связывание ионов Са2+ разветвленной сетью трубочек и цистерн сарко-плазматической сети не является простой адсорбцией. Это активный физиологический процесс, который осуществляется за счет энергии, освобождающейся при расщеплении АТФ Са2+-зависимой АТФазой саркоплазматической сети. В целом такой механизм получил название «кальциевая помпа» по аналогии с хорошо известным в физиологии натриевым насосом.
Возможность пребывания живой мышцы в расслабленном состоянии при наличии в ней достаточно высокой концентрации АТФ объясняется снижением в результате действия кальциевой помпы концентрации ионов Са2+ в среде, окружающей миофибриллы, ниже того предела, при котором еще возможны проявление АТФазной активности и сократимость акто-миозиновых структур волокна.
Быстрое сокращение мышечного волокна при его раздражении от нерва (или электрическим током) является результатом внезапного изменения проницаемости мембран и как следствие выхода из цистерн и трубочек саркоплазматической сети и Т-системы некоторого количества ионов Са2+ в саркоплазму.
Почему ионы кальция все же оказывают такое сильное влияние на сократительные свойства мышцы? Почему в присутствии этих ионов мышца может сокращаться? А в их отсутствии сокращения не происходит?
Дело в том, что, как мы помним, к актиновым филаментам присоединен топонин-тропомиозиновый комплекс, который в отсутствии ионов кальция “закрывает” активные центры актина и, по сути, не дает возможности “головкам” миозина присоединиться.
При передаче нервного импульса к поверхности сарколеммы происходит “выплеск” ионов кальция в саркоплазму. Эти ионы кальция связываются с тропонином, при этом происходят конформационные изменения, которые, по-видимому, приводят к сдвигу всего тропонин-тропомиозинового стержня и деблокировке активных центров актина, способных взаимодействовать с миозином.
Наконец, если возбуждение прекращается, содержание ионов Са2+ в саркоплазме снижается (кальциевая помпа), то циклы прикрепление–освобождение прекращаются, т.е. «головки» миозиновых нитей перестают прикрепляться к актиновым нитям. В присутствии АТФ мышца расслабляется и ее длина достигает исходной. Если прекращается поступление АТФ (аноксия, отравление дыхательными ядами или смерть), то мышца переходит в состояние окоченения. Почти все поперечные мостики толстых (миозиновых) нитей присоединены при этом к тонким актиновым нитям, следствием чего и является полная неподвижность мышцы.
Небольшой ролик по теме:
Стоить отметить важный факт. В ходе ресинтеза АТФ (в ходе анаэробного гликолиза, о котором мы поговорим позднее) образуется большое количество молочной кислоты, которая закисляет мышцу. Ионы водорода конкурируют с ионами кальция на активных центрах, что приводит к прекращению сокращения мышцы.
Двигательная единица.

Двигательная единица (ДЕ) состоит из клеточного тела α-мотонейрона переднего рога спинного мозга, его аксона (который проходит по спинномозговому, а затем — по двигательному нерву, входя в мышцу, где он разветвляется на множество мышечных ветвей), и многочисленные концевые двигательные пластинки, где каждая нервная веточка заканчивается на единственном мышечном волокне (т.е. клетке).
ДЕ включает все мышечные волокна, иннервируемые одним мотонейроном. Любое мышечное волокно в норме получает нервное обеспечение только из одной концевой двигательной пластинки и потому только из одного мотонейрона.
Количество ДЕ остается неизменным у человека, а так же количество МВ практически не изменяется в ходе тренировок.
Однако с помощью тренировочного процесса можно добиться гиперплазии (рост количества элементов) многих органелл: миофибрилл, митохондрий, СПР, глобул гликогена и т.д.
Таким образом, один мотонейрон включает в работу несколько мышечных волокон.
Мотонейроны различаются возбудимости (скорости включения в работу). Так медленные (низкопороговые – низкий порог включения в импульсную активность), мотонейроны (они, кстати, и более мелкие по размерам) входят в т.н. НИЗКОПОРОГОВЫЕ ДЕ, соответственно, быстрым (большим, высокопороговым) мотонейронам соответствуют ВЫСОКОПОРОГОВЫЕ ДЕ.
Причем здесь какой-то “порог”?
Дело в том, что при увеличении нагрузки сначала включаются низкопороговые ДЕ, дальнейшее увеличение нагрузки приводит к иннервированию высокопороговых ДЕ.
Это т.н. “Принцип величины” Хеннемана.
С другой стороны, низкопороговым ДЕ соответствуют т.н. окислительные (красные) МВ – ОМВ, а высокопороговым – гликолитические (белые) МВ – ГМВ (о классификации МВ мы поговорим чуть позже).
Если быть максимально точным, то все же в самом начале работы мышцы на некоторое время задействуются высокопороговые МВ (посредством больших (быстрых) мотонейронов) чтобы увеличить скорость нарастания напряжения мышц («градиент силы») или сообщить двигаемой части тела необходимое ускорение. Затем, если нагрузка незначительна, высопороговые ДЕ выключаются из работы и используются только низкопороговые ДЕ.
Скорость проведения импульса по аксону в прямой связи с диаметром аксона. Соответственно в большинстве случаев скорость проведения импульса у медленных мотонейронов ниже, чем у быстрых, т.к. медленные (малые) мотонейроны имеют относительно тонкий аксон, а большие (быстрые) мотонейроны – толстый аксон.
Частота импульсации медленных мотонейронов ниже, чем у быстрых. Это касается как начальной (пороговой), так и максимальной частоты импульсации. Соответственно частотный диапозон у быстрых мотонейронов значительно выше.
Следовательно, за счет изменения частоты импульсации мотонейрона напряжение мышечных волокон быстрой ДЕ может изменяться в значительно большей степени, чем напряжение мышечных волокон медленной ДЕ.
Утомляемость или обратный показатель – выносливость также различается у двух типов мотонейронов. Медленные мотонейроны способны поддерживать длительный разряд без заметного снижения частоты импульсаций на протяжении десятков минут. Поэтому их называют неутомляемыми мотонейронами.
Быстрые мотонейроны относятся к утомляемым, так как они не способны к длительному поддержанию высокочастотных импульсаций.
Что это нам дает? Понимание того, что в начальный момент времени, до достижении некоторого усилия, работают эти самые ОМВ, они наиболее выносливые и, в принципе, могут работать бесконечно долго без закисления. Время работы фактически лимитируется количеством питательных веществ.
После того как усилие достигает определенного предела начинают включаться в работу ГМВ. Они не такие выносливые как ОМВ, по мере работы в них накапливаются продукты распада питательных веществ, которые способствуют прекращению работы мышцы в целом. Кроме того, как было замечено выше, ГМВ интервируются быстрыми мотонейронами, которые быстро утомляются и не могут поддерживать необходимую высокую частоту пульсации.
Кстати, умение включать в работу одновременно большое количество ДЕ, как низкопороговых, так и высокопороговых, определяет максимальную силу данной мышцы (это достигается тренировками на максимальную силу – работа с предельными весами, минимальное кол-во повторений).
Это умение очень важно для болдыринга.
Классификация МВ.
Существует несколько классификаций мышц:
1. Красные/Белые
Красные МВ.
Эти волокна окружены огромным количеством капилляров, а названием своим обязаны белку миоглобину, повышенное содержание которого и придает волокну красный цвет.
Белые МВ.
В таких волокнах, наоборот, миоглобина очень мало. Поэтому они и белые.
2. Медленные/Быстрые.
Быстрые и медленные мышечные волокна содержат изоферменты миозина , которые расщепляют АТФ с разной максимальной скоростью; этому соответствует различная максимальная скорость рабочего цикла поперечных мостиков и, следовательно, укорочения волокна. Высокая АТФазная активность миозина свойственна быстрым волокнам, более низкая – медленным волокнам.
АТФазная активность миозина – наследуемый фактор и не подвергается тренировке.
3. Окислительные/Гликолитические.
Этот подход к классификации волокон скелетных мышц основан на различиях ферментативных механизмов синтеза АТФ. В некоторых волокнах много митохондрий , и, следовательно, обеспечивается высокий уровень окислительного фосфорилирования; это окислительные волокна. Количество образующейся в них АТФ зависит от снабжения мышцы кровью, с которой поступают молекулы кислорода и богатых энергией соединений. Волокна этого типа окружены многочисленными капиллярами. Кроме того, в них присутствует связывающий кислород белок – миоглобин , увеличивающий скорость диффузии кислорода, а также выполняющий роль кратковременного кислородного депо в мышечной ткани. Благодаря значительному содержанию миоглобина окислительные волокна окрашены в темно-красный цвет; их часто называют красными мышечными волокнами.
В гликолитических волокнах , наоборот, мало митохондрий, но высокое содержание ферментов гликолиза и большие запасы гликогена . Эти волокна окружены относительно небольшим числом капилляров, и миоглобина в их ткани немного, что соответствует ограниченному использованию кислорода. Вследствие недостатка миоглобина гликолитические волокна выглядят светлыми и получили название белых мышечных волокон.
На основании двух рассмотренных характеристик (скорость укорочения и тип метаболизма) можно выделить три типа волокон скелетных мышц.
– Медленные ОМВ (тип I) – низкая активность миозиновой АТФазы и высокая окислительная способность.
– Быстрые ОМВ (тип IIа) – высокая активность миозиновой АТФазы и высокая окислительная способность.
– Быстрые ГМВ (тип IIб) – высокая активность миозиновой АТФазы и высокая гликолитическая способность.
Отмечу, что не обнаружен четвертый теоретически возможный вариант – медленные гликолитические волокна.
Волокна варьируются не только по своим биохимическим особенностям, но и по размерам: у гликолитических волокон диаметр существенно больше, чем у оксидативных. Это сказывается на величине развиваемого ими напряжения. Число толстых и тонких филаментов на единицу площади поперечного сечения примерно одинаково для всех типов скелетных мышечных волокон. Таким образом, чем значительнее диаметр волокна, тем большее число параллельно задействованных толстых и тонких филаментов участвует в генерировании силы и тем больше, наконец, максимальное напряжение мышечного волокна. Отсюда следует, что гликолитическое волокно, имеющее больший диаметр, развивает в среднем более значительное напряжение по сравнению с напряжением окислительного волокна .
Кроме того, рассмотренные три типа мышечных волокон характеризуются разной устойчивостью к утомлению . Быстрые ГМВ утомляются через короткое время, тогда как медленные ОМВ волокна очень выносливы, что позволяет им длительно поддерживать сократительную активность практически при постоянном уровне напряжения. Быстрые ОМВ занимают промежуточное место по способности противостоять развитию утомления.
Несмотря на то, что АТФазная активность наследуется, с помощью тренировок можно повысить окислительный потенциал ГМВ т.е. сделать их более выносливыми. Для этого необходимо увеличить количество митохондрий в ГМВ.
Для любого вида спорта необходимо понимание какие типы мышц нужны. Например, в длительном, марафонском, беге важную роль будут играть ОМВ – нужно бежать долго и без закисления. Тоже, вероятно, можно предположить, важно и для лазания (мышцы предплечий в основном) по длинным маршрутам.
Для спринтера важно развить максимальное ускорение и здесь ему помогут ГМВ. Для бегунов на средние дистанции важен некоторый баланс между ОМВ и ГМВ – большую часть дистанции спортсмен пробегает на ОМВ и на финише дает ускорение с помощью ГМВ.
Для скалолазания, вероятно, так же нужен некий баланс. Если у вас предплечья полностью из ГМВ – вы пролезете один жесткий болдеринг или одну сложную трассу и все. Накопившейся молочной кислоте деваться будет некуда – нет рядом ОМВ, которые могли бы быстро утилизировать ионы водорода. Если же у вас только ОМВ, вероятно, вы не сможете развить своего максимального усилия.
Хотя часто в максимальных усилиях нет необходимости, часто дело в другом – не техничное лазание, страх, психологическая неподготовленность.
Поэтому я склонен предположить, что для мышц предплечий (часто это самое слабое звено) важно быть в большей степени окислительными. Но, конечно, какая-то часть волокон должна быть гликолитической для развития максимального усилия.
Итого. Что мы имеем? Попробую коротко.
Мышцы представляют собой пучки мышечных волокон. Каждое мышечное волокно – большая многоядерная клетка. Большая часть этой клетки заполнена миофибриллами, белковыми нитевидными образованиями. Эти миофибриллы и отвечают за сокращение.
Они состоят в основном из двух белков – актина и миозина.
Кроме миофибрилл в каждом мышечном волокне еще много всяких органелл – ядра, митохондрии, некоторое количество гликогена и липидов(питательные вещества), креатинфосфат и т.д. Так же внутри такой клетки есть очень важное образование – саркоплазматический ретикулум – это такая хитрая система трубочек, канальцев, цистерн, в которых содержится большое количество ионов кальция.
Как мышцы сокращаются?
К каждому МВ подходят нервные окончания из спинного мозга. Когда из мозга поступает сигнал на поверхность мышечного волокна, этот импульс заставляет саркоплазматический ретикулум выплеснуть ионы кальция. Это приводит к тому, что актин и миозин начинают взаимодействовать друг с другом и начинается мышечное сокращение – ионы кальция “помогают” в этом процессе.
Когда импульсы перестают поступать, включается т.н. “кальциевый насос”, который откачивает ионы кальция обратно в ретикулум. Ионов кальция нет – сокращения нет. Происходит расслабление мышцы.
Этот процесс (откачивание) происходит при затрате энергии. Если организм мертв, энергия не вырабатывается, кальциевый насос не работает, мышца расслабится не может – трупное окоченение.
В конкретном мышечном волокне может быть много митохондрий или мало. Соответственно имеем окислительные МВ или гликолитические МВ соответственно. Конкретная мышца состоит из разных типов МВ. Если мышца много и часто работает – там больше ОМВ, если она работает мало – мышцы в основном гликолитические.
Зачем нужны митохондрии?
В них происходит т.н. аэробный гликолиз (питательные вещества разрушаются наиболее эффективно, выделяется большое количество энергии. Конечными продуктами являются углекислый газ и вода). Следовательно, если в МВ много митохондрий (т.е. они являются окислительными), то работа таких мышц наиболее эффективна с точки зрения утомляемости. Они могут работать очень долго, хотя и не могут развить максимальную силу.
Если митохондрий мало – это гликолитические МВ. В них идет в основном анаэробный гликолиз. В ходе этого типа гликолиза разрушение питательных веществ идет до молочной кислоты. Энергии, по сравнению аэробным путем, выделяется меньше.
Ионы водорода (из молочной кислоты) конкурируют с ионами кальция и мешают мышце сокращаться.
Такие МВ могут развить большую силу, но они быстро утомляемые.
С помощью тренировок можно гликолитические МВ сделать окислительные.
Для скалолазания важны и те и другие.
“Принцип величины” Хеннемана.
Двигательная единица – комплекс состоящий из клеточного тела α-мотонейрона переднего рога спинного мозга, его аксона (который проходит по спинномозговому, а затем — по двигательному нерву, входя в мышцу, где он разветвляется на множество мышечных ветвей), и многочисленные концевые двигательные пластинки, где каждая нервная веточка заканчивается на единственном мышечном волокне
При увеличении нагрузки сначала включаются низкопороговые двигательные единицы (чаще всего окислительные МВ), дальнейшее увеличение нагрузки приводит к иннервированию высокопороговых ДЕ (чаще всего гликолитические МВ).
Короче говоря, сначала, при низкой нагрузке, в работу включаются окислительные МВ и они могут работать очень долго. Например, ходьба.
Затем, при увеличении, нагрузки (например, переходим на бег или начинаем идти в горку) подключаются гликолитические. Но т.к. работа таких МВ обеспечивается преимущественно анаэробным гликолизом, то в результате накапливается большое количество ионов водорода, что в итоге приводит к закислению мышцы.
Причем тут скалолазание?
Для болдыринга, вероятно, нужны преимущественно ГМВ, хотя если будут лишь такие волокна то один раз закислившись, нужно будет много времени прежде чем ионы водорода будут утилизированы.
Для длинных трасс мышцы предплечий, предполагаю, должны иметь высокую долю ОМВ. ГМВ в этом случае тоже нужны – для прохождения сложных ключей.
Нужно понимать, что пока вы лезете простые участки – работают в основном ОМВ, когда увеличивается сложность (и/или становится страшно, появляется неуверенность) начинают подключаться высокопороговые ДЕ – ГМВ. И, конечно, это начало конца. Ионы водорода начинают накапливаться, буферная емкость системы постепенно переполнится и мышца перестанет работать.
Что можно сделать?
Приходить на тренировки 🙂
Ну вот и все что я хотел рассказать поэтому поводу.
В следующем посте я попытаюсь рассказать все что знаю про энергообеспечение мышцы.
 Kalymnos, 2016 Edition.
Kalymnos, 2016 Edition. Неожиданный апрельский Крым.
Неожиданный апрельский Крым. Открытая закрытая Турция.
Открытая закрытая Турция. Игры в мафию. Конец игры.
Игры в мафию. Конец игры. Игры в мафию. Игра продолжается.
Игры в мафию. Игра продолжается. Игры в мафию. День 2. Первый дождь.
Игры в мафию. День 2. Первый дождь. Итальянские приключения.
Итальянские приключения. Игры в мафию.
Игры в мафию.

